Reja:
1.Dasturlashgan hujayra o’limi va uni
xususiyatlari
2.Dasturlashgan hujayra o’limining
retseptorlari
3.Apoptozning funksional malekulalar
guruhi
4. VICH bilan kasallangan hujayralarda
apoptoz
Barcha ko`p hujayrali organizimlarda
shuning dek odamda ham hujayra o`limi
dasturi mavjuddir.
Tashqi yoki ichki omillar ta’siri natijasida
o’z-o’zini yo’qotish dasturi faollashgan
holatda apoptoz nomli hujayra o’limi ro’y
beradi.Apoptoz kuzgi barg to’kilishini
misol qilishimiz mumkin.Apoptoz
bu,umumbiologik xodisa bo’lib qari yoki
kasal hujayralardan qutilishga imkon
beradi.
Bunday hujayralardan qutilishga sabab ,1
chidan tirik va o’lik hujayralar o’rtasidagi
tenglikni saqlash,2 chidan mutatsiyalar
hujayralar rivojlanishining nasliy dasturini
buzmasligi uchun.Hujayra o’limi 2 ta
xususiyatga ega;bir tomondan u gomeostozni
saqlashga xizmat qilsa,boshqa tomondan
organizm va uning avlodini xavfdan himoya
qiladi.Hujayraning dasturlashgan o’limi
organlar shakllanayotganda embriogenezning
boshlang’ich bosqichlarida aniqlanadi.
Ya’ni bir to’qima boshqasiga almashganda,
vaqtinchalik a’zolar rezorbsiyasiga va boshqa
jarayonlarda,yetuk organizmlarda apoptoz mitozga
komplenentar bolib, hujayra populyatsiyasini
boshqarishda ishtirok etadi, ayniqsa organizm
patogenlar bilan zararlanganda vaqtincha
limfositlar soni ko’payadi. Shularni yo’qotishga
xizmat qiladi.Shu bilan bir vaqtda apoptoz hujayra
illiminasiyasi, ya’ni genetik buzilgan yoki
o’smaga aylanayotgan hujayralarni yo’qotish
uchun kerak.
Apoptoz boshqaruvining ishdan chiqishi
organizmda patologik o’zgarishlarga olib keladi.Bu
holat antiapoptoz faollikni to’suvchi oqsillarni
kodlovchi genlarga ega viruslar ta’sirida sodir
bo’ladi.Shuning uchun apoptozning hujayraviy va
molekulyar xususiyatlari onkogenez va virusli
patogenez bilan bog’langan va hujayra bo’linish
boshqaruvining nozik mexanizmini o’rganishga
aloqadordir.So’nggi yillarda hujayraning
dasturlashtirilgan o’limi yaxshi o’rganilmoqda.
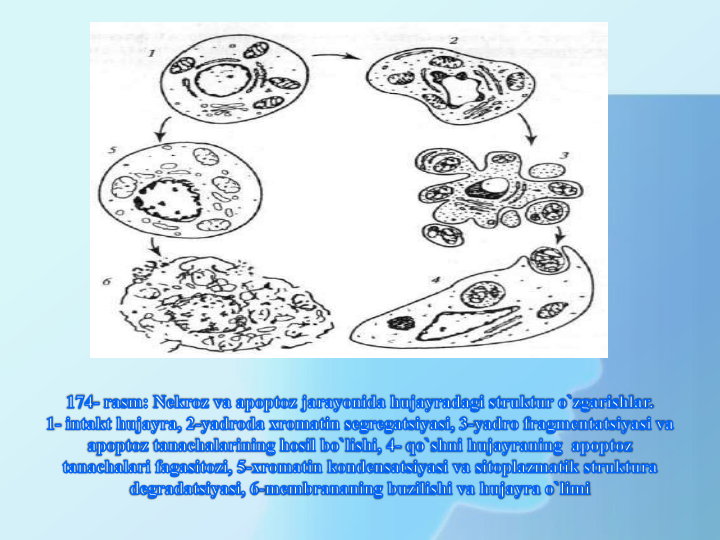
Sitologik jihatdan apoptoz hujayra o’limining
boshqa shakli bo’lish nekrozdan farq qiladi,u og’ir va
birdaniga sodir bo’lgan hujayra zararlanish natijasida
sodir bo’ladigan metabolitik halokat sifatida qaraladi.
Hujayraning nekrotik o’limi fiziologik yoki kimyoviy
jarohatlar chaqiradi. Nekrozda hujayra
shishib,sitoplazmatik va hujayra ichi membranalari
sturukturasi o’zgaradi, bunda lizosoma fermentlari
sturukturasi o’zgaradi, bunda lizosoma fermentlari
ajralib chiqadi va hujayra lizisga uchraydi.
Nekroz apoptozdan farqli
yalig’lanish jarayoni bilan kechadi.
Nekrozdan farqli apoptoz xromatik
destruksiyasida kuzatiladi, bunda
hujayra hajmi kichrayib, yadro bo’linib
ketadi. Apoptoz hujayra bo’linib ketish
va apoptoz tanalarning hosil bo’lishi
bilan tugaydi, ular keyin tezda
makrofaglar tomonidan parchalanib
tashlanadilar.
Nekrozdan
farqli
apoptoz
xromatik
destruksiyasida
kuzatiladi,bunda
xujayra
xujayra xajmi kichrayib, yadro’ bo’linib ketadi.
Apoptoz xujayra bo’linib ketish va apoptoz
tanalarning hosil bo’lishi bilan tugaydi ular
keyin
tezda
makrofaglar
tomonidan
parchalanib
tashlanadilar.
Apoptoz
uchun
DNKning
fermentative
nukleosomalararo
fragmentatsiyasi xosdir, bu jarayon hujayra
membranasini parchalanishini oldidan sodir
bo’ladi.
Malekulyar jihatdan apoptoz ko’p
bosqichli jarayondir,u ma’lum signal
qabul qilishdan boshlanib,litik fermentlar
ta’sirida hujayra molekulalari va
sturukturalarini buzilishi bilan
tugaydi.Signal ma’lum retseptorlar bilan
qabul qilinadi va vositachi molekulalar
orqali hujayra yadrosiga yetkaziladi,u
yerda apoptoz rivojlanishini
determenlovchi genlar ekspressiyasi sodir
bo’ladi.
Hozirgi paytda apoptoz jarayoniga oid 4ta
asos funksional molekulalar guruhi aniqlangan;
proteolitik
fermentlar
(kaspazalar);
faolligini
boshqaruvchi adapter oqsillar; o’smalar nekro’zi
omillar retseptori oilasi oqsillari (TNF) va BCL-
2 oilasi oqsillari. Hayvon va odam hujayralarida
apoptoz kaspazalar bilan bog’liq, ular sistein
proteinazalar bolib’ asparagin kislota qoldig’ini
parchalovchi oqsillardir. Sutemizuvchilarda 14 ta
kaspaza aniqlangan.
Ular
oqsil
polipeptid
zanjirida
tetrapeptit
bloklarda
aniqlab,asp
qoldig’ining
karboksil
guruhi
hosil
qilgan
peptid
bog’ini
parchalaydi.
Kaspazalar
juda
past
faolikka
ega
profermentlar
shaklida
sintezlanadi.
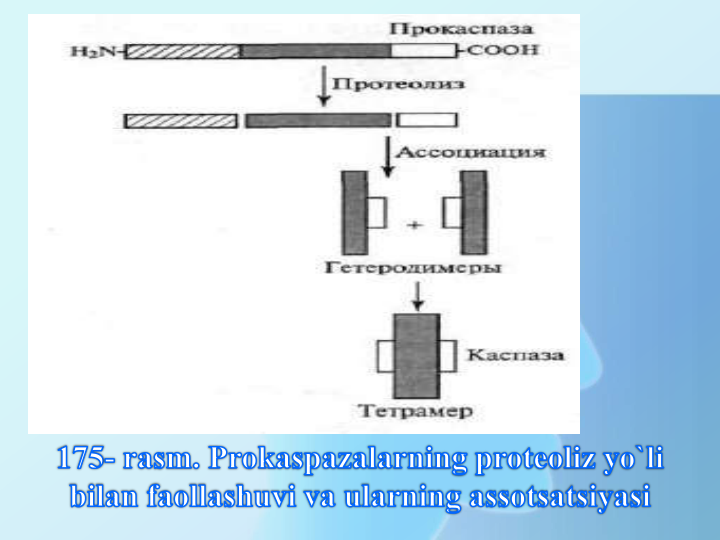
Yetuk kaspazalar tetramerlar bo’lib,2 ta
20
KDa
va
10
KDa
li
2
tadan
subbirliklardan
tuzilgan.Faol
kaspazalar
proteolitik
o’zini-o’zi
parchalash
yoki
kesishuvchi
parchalanish orqali hosil bo’ladi.
Bu jarayon davomida prokaspazadan
N-uchli domen ajraladi,qolgan
molekula qismi katta va kichik
subbirlikka ajraladi. So’ngra ular
geterodimer va tetramer hosil qiladi.
Kaspazalarni initsirlovchi 8 va 9
faollashuvchi yoki effector 3, 6, 7,
turlari farq qilinadi. Masalan, 8
kaspaza 3 prokaspazani faollashtiradi,
undan faol effector 3 kaspaza hosil
bo’ladi.
Effektor kaspazaning faollashuvi apoptozga qaytmas
tus beradi.Kaspazalar,ayniqsa effector kaspazalar hujayra
hayoti
uchun
muhim
bo’lgan
oqsillar,ya’ni
DNK
reparatsiyasi
fermentlari,
p-53
oqsili
ingibitori,C
proteinkinaza
va
boshqalarni
parchalab
nofaollashtiradi.Kaspazalarning o’ljalaridan biri
DNK
reparatsiyasi va transkripsiyasi bilan bog’liq polimeraza
fermentidir. Kaspazalar faollashtiradigan CAD DNK-aza
odatda DFF ingibitori bilan kompleks nofaol xolatda
bo’ladi.Apoptoz jarayonida DFF kaspazalar tamonidan
parchalanib, faol CAD endonukleazani ozod qiladi, u
linker
DNK
atrofida
xromatinning
nukleosom
uzulishlarini
amalga
oshiradi.
Natijada
nuklesom
bo’lagiga teng DNK bo’laklari xosil bo’ladi.
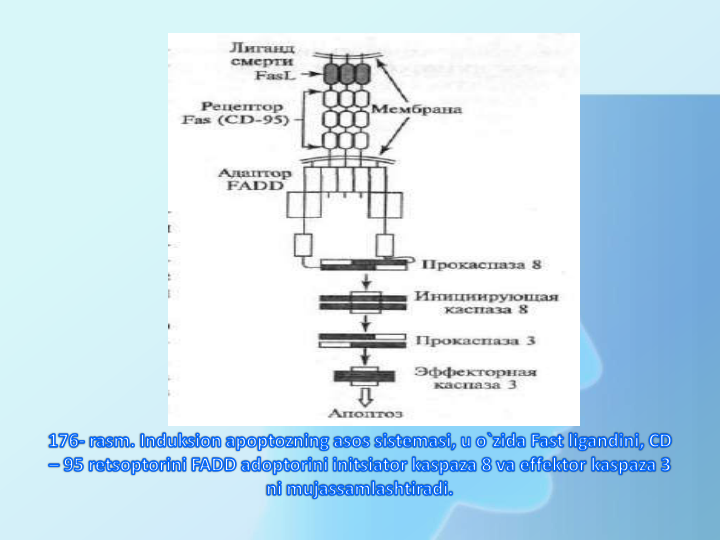
Hujayrada apaptozning aloxida retseptorlari bo’lib,
ular tegishli ligandlardan signal qabul qiladi va hujayra
o’limi dasturini stimullaydi. Masalan, Fas retseptori
ta’lluqli ligand bilan ta’sirlashib viruslar bilan kasallangan
hujayrada o’lim dasturini faollaydi, Fas L o’sma nekroz
omili
TNF
ligandlar
oilasiga
tegishli.
TNF
retsptorlari
transmembrana
oqsili
bo’lib,
o’zining hujayradan tashqaridagi domenlari ligandlarning,
trimer
malekulalarini
apoptoz
induktorlari
bilan
tasirlashadi,
ularning
sitoplazmatik
qisimlar
apoptoz
asosning hujayra ichi ishtirokchilari bilan ta’sirlashishga
xizmat qiladi.
Apoptozda spetsifik genlar ishtirokining isboti 1-
chi marta Caenorhabditis elegans nematodasida
dasturlashgan hujayra o’limini o’rganish mobaynida
olingan ularda 1090 tadan 131 somatik hujayralar
rivojlanishining ma’lum bosqichida halok
bo’ladi,chuvalchanglarning bu turida hujayra o’limi
uchun ced 3va ced 4 genlari kerak, ced 9 geni o’limni
to’xtatuvchi oqsilni kodlaydi. Sutemizuvchilarda
bunday gen bcl-2 dir, bu oilaga struktur –qondosh
oqsillarning 2 turini kodlovchi genlar kiradi.
Birinchi
turga sutemizuvchilarning apoptozga
to’sqinlik qiluvchi, shuningdek, viruslarning
bcl-2
kiradi,
u
bcl-2ga
o’xshash
1lamchi
struktura
qismlarini
tutadi,
2
chi
turiga
apoptozni
kuchaytiruvchi oqsillar kiradi. Bcl -2 proapoptoz
oqsillarning faolligi posttranslyatsion modifikatsiya
bosqichida
boshqariladi:
fosforlar
va
cheklangan
proteoliz
orqali.
Apoptoz induktorlari ta’siriga uchragan hujayralarda
ularning ichki membrana
yirik teshikchalaridagi
metoxondriya
matriksning
ishishi
va
tashqi
membrananing
uzilishi
kiradi.





” mavzusidagi")










;
faolligini
boshqaruvchi adapter oqsillar; o’smalar nekro’zi
omillar retseptori oilasi oqsillari (TNF) va BCL-
2 oilasi oqsillari. Hayvon va odam hujayralarida
apoptoz kaspazalar bilan bog’liq, ular sistein
proteinazalar bolib’ asparagin kislota qoldig’ini
parchalovchi oqsillardir. Sutemizuvchilarda 14 ta
kaspaza aniqlangan.")